1 Einleitung
Seit ihrer Entdeckung im 19. Jahrhundert erfreuen die wunderschön anzusehenden Zellen der Zieralgengattung Micrasterias jede Betrachterin und jeden Betrachter. Da die meisten Arten relativ große und flach gewachsene Zellen haben, gestatten sie meist einen guten Einblick in Zelldetails wie Chloroplasten, Zellkerne und sogar sehr kleine Zellorganellen wie Mitochondrien und Dictyosomen. Manchmal werden auch die Gallertschichten sichtbar, welche den Zellwänden aufliegen. Bei höherer Vergrößerung sind zudem oft die feinen Poren erkennbar, durch welche die Zelle die Gallerte auf die äußere Seite der Zellwand transportieren kann. Dass diese Wände mehrschichtig aufgebaut sind, können wir zwar in der Literatur lesen, aber lichtmikroskopisch nicht beobachten. Hierbei half die Elektronenmikroskopie.
2 Probenvorbereitung
Da die Abmessungen der zu untersuchenden Strukturen deutlich unterhalb der lichtmikroskopischen Auflösungsgrenze liegen, kam ein Rasterelektronenmikroskop (REM) mit Schottky-Feldemissions-Kathode (Tescan Clara) zum Einsatz. Durch Verwendung einer Schottky-Kathode kann die Auflösung im Vergleich zu REMs mit thermischen Emittern um ungefähr eine Größenordnung verbessert werden. Gleichzeitig werden diese Auflösungen bei deutlich verringerten Beschleunigungsspannungen ermöglicht, was für die Beobachtung empfindlicher biologischer Proben essenziell ist (Wanner, 2023).

Abbildung 1: Verwendete Gerätschaften für die Probenpräparation und -analyse: Kritisch-Punkt-Trockner (Leica EM CPD300), Sputter Coater (Cressington 108 Auto) und Rasterelektronenmikroskop (Tescan Clara).
Da die Proben im Hochvakuum untersucht werden, müssen die Zieralgen vorab getrocknet werden. Bei Trocknung an der Luft entstehen Schrumpfungsartefakte, deren Ursache in der Kapillarwirkung des verdampfenden Wassers liegt: Die gasförmige Phase des Wassers wird über die Luft abtransportiert, während die flüssige Phase im Laufe des Trocknungsprozesses allmählich verschwindet. Durch Kapillareffekte werden die festen Zellbestandteile zusammengezogen und die ursprüngliche Morphologie des Gewebes wird zerstört.
Um diesen Vorgang zu unterbinden, müssen also Phasenübergänge vermieden werden. Dies wird beim Kritisch-Punkt-Trocknen (Critical Point Drying – CPD) dadurch erreicht, dass flüssiges CO2 zum Einsatz kommt. Das Wasser in der Probe wird zuerst durch Spülen mit wasserfreiem Ethanol entfernt. Das Ethanol wird anschließend bei Temperaturen zwischen 15 °C und 20 °C unter Druck (ca. 50 bar) durch flüssiges CO2 ersetzt. In weiterer Folge wird der kritische Punkt von CO2 durch Erhöhung von Druck und Temperatur (siehe orange Linie in Abbildung 2) umgangen. Druck und Temperatur werden langsam abgesenkt, damit das jetzt gasförmige CO2 aus der Probe entweichen kann. Am Ende steht eine getrocknete Probe, deren flüssige Bestandteile ohne Überschreiten einer Phasengrenze entfernt worden sind. Die Kritisch-Punkt-Trocknung ermöglicht somit eine artefaktarme Präparation von Einzellern für die Beobachtung im REM.
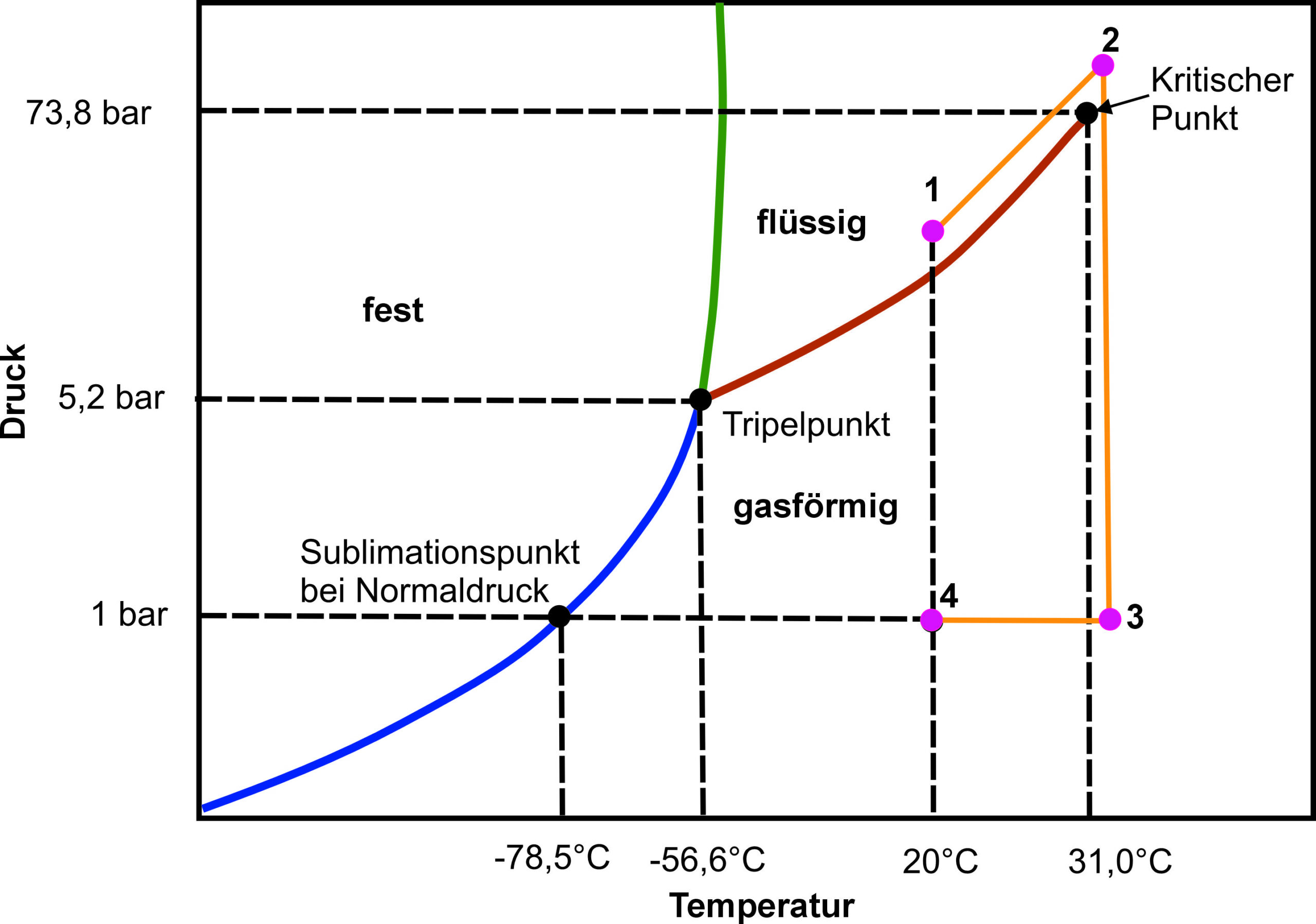
Abbildung 2: Phasendiagramm von CO2. Die orange Linie (Punkt 1-4) zeigt schematisch den Druck-Temperatur-Verlauf beim Kritisch-Punkt-Trocknen.
3 Beobachtung im REM
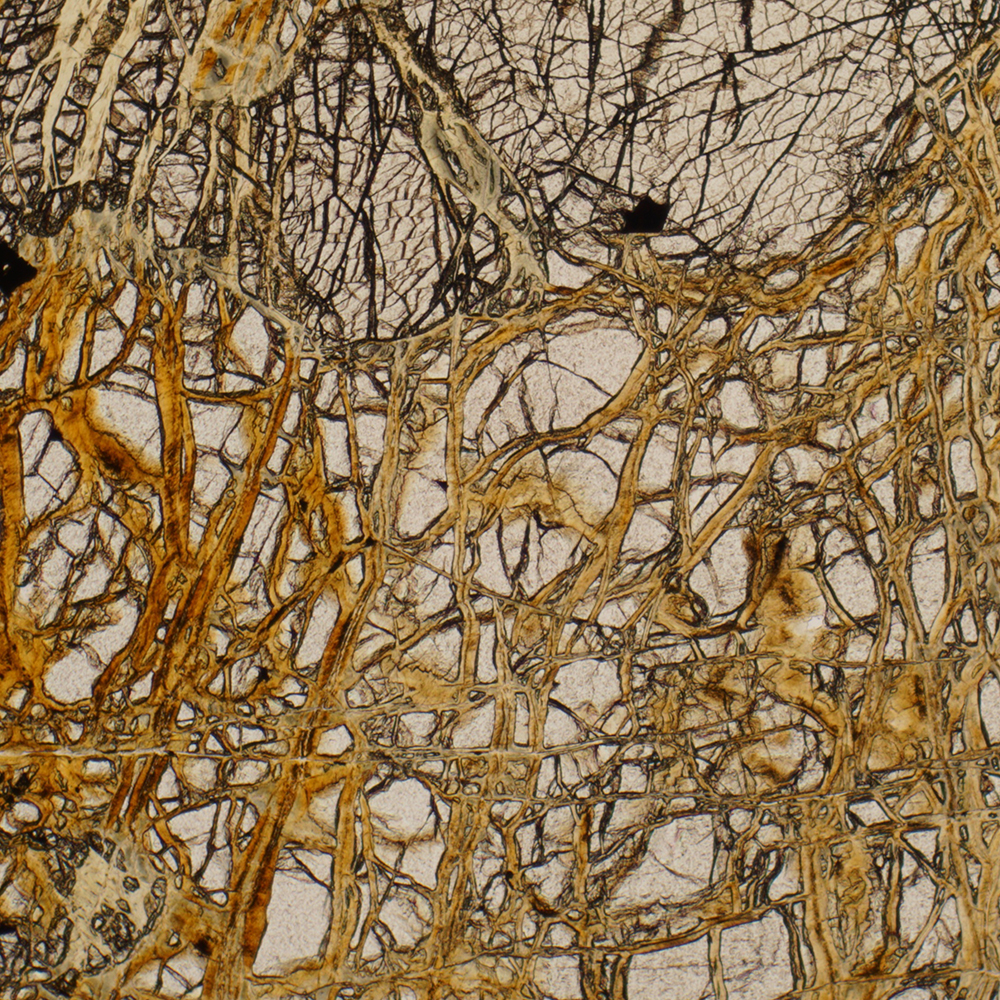
Nach der CPD-Verarbeitung einer Zieralgenprobe aus dem Moor Schwemm bei Walchsee in Tirol wurden die daraus gewonnenen Zellen im Sputter Coater mit einer dünnen Goldschicht besputtert und im REM beobachtet. Die Beschleunigungsspannung lag zwischen 1 und 2 kV, um eine Schädigung der empfindlichen Fibrillen möglichst zu vermeiden. Im Endergebnis konnten das Netz der Cellulose-Mikrofibrillen der Primärwand bei Micrasterias fimbriata sowie die Anschwellungen der Porenapparate in der Sekundärwand präzise dargestellt werden (Abbildung 3).


Abbildung 3: Micrasterias fimbriata aus dem Moor „Schwemm“ bei Walchsee in Tirol (Österreich). Bereits im Übersichtsbild wird ein netzartiger Überzug sichtbar.
3.1 Aufbau der Zellwand
Bei der Zellteilung von Zieralgen, aber auch bei Gefäßpflanzen, entsteht bei jeder neu gebildeten Zelle zunächst die sogenannte Primärwand. Sie ist sehr dünn (50–100 nm) und weich, aber flexibel und schützt die heranwachsende Zelle. Die Primärwände bestehen aus Zellwandgrundsubstanz (Pektin, Hemizellulosen), besitzen aber auch schon ein dünnes Netz aus Cellulose-Mikrofibrillen. Haben sich der Zellkern sowie die Chloroplasten und die Mitochondrien geteilt und sind die Teilungsprodukte komplett in die neue Zelle eingewandert, beginnt diese, die Sekundärwand aufzubauen, welche die für die Art typische Skulptur und Ornamentierung bekommt sowie die benötigte Stabilität liefert (Kleinig & Maier, 1999). Von Cosmarium-Arten ist bekannt, dass die Primärwand nach der abgeschlossenen Zellteilung abgestoßen wird, bei Micrasterias umhüllt sie die Gallertschicht aus Mucopolysacchariden, die der widerstandsfähigen Sekundärwand aufliegt. Diese in den Dictyosomen der Zelle produzierte Gallerte wird über die Porenapparate nach außen transportiert und schützt die Zelle zusätzlich (Abbildung 4).
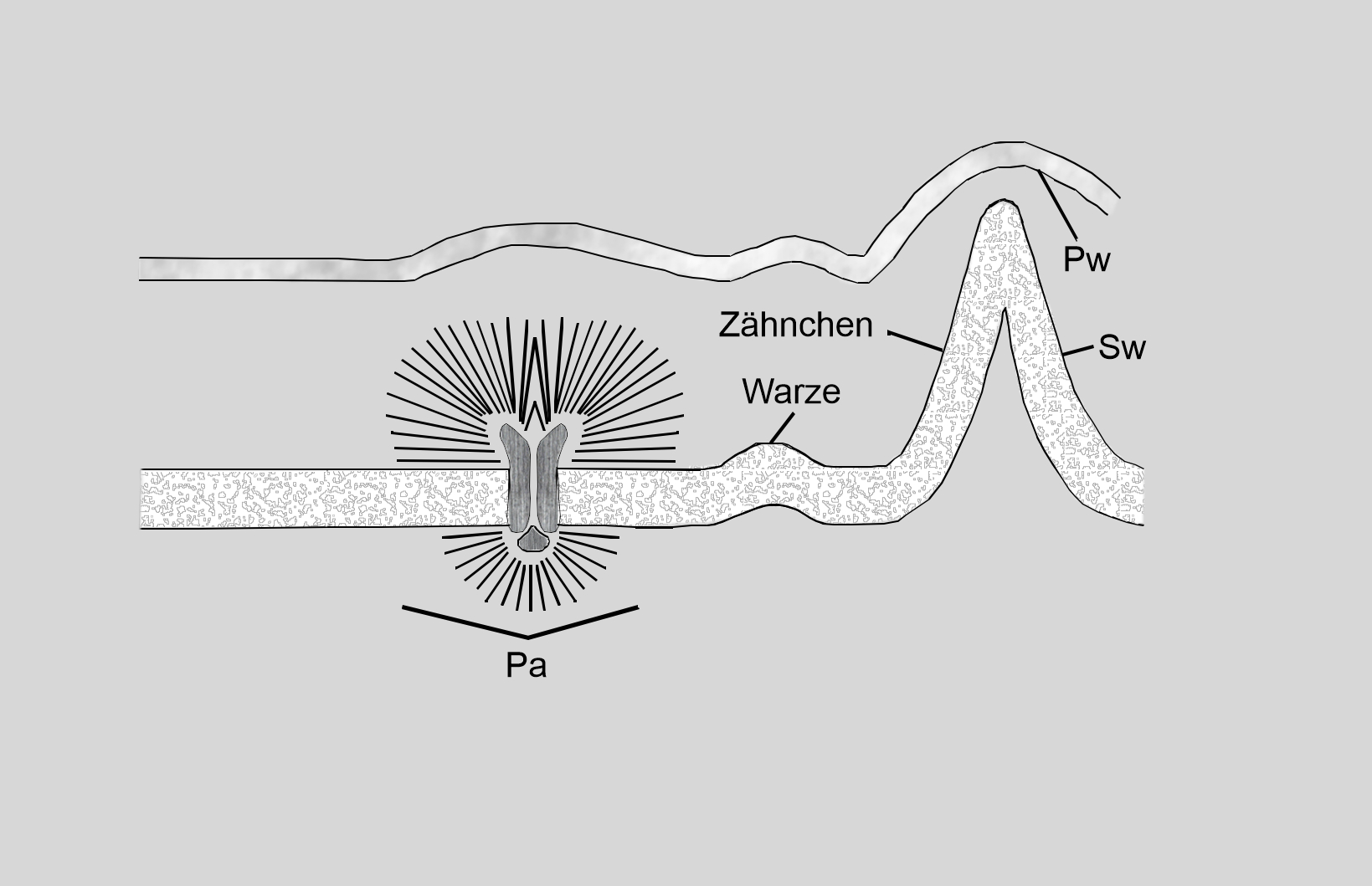
Abbildung 4: Schemazeichnung eines Querschnitts durch eine Desmidiaceen-Zellwand vom Cosmarium-Typ, zu welchem auch die Zellwände von Micrasterias gehören. Pw = Primärwand, Sw = Sekundärwand, Pa = Porenapparat. Nach Brook (1981), verändert.
3.2 Die Cellulose-Mikrofibrillen der Primärwand
Die Abbildung 5 sowie die Vergrößerungsreihe in Abbildung 6a–c zeigen eindrucksvoll das Maschenwerk der Cellulose-Mikrofibrillen der Primärwand, die exemplarisch mit Pfeilen markierten Höcker stammen von den Porenapparaten in der darunterliegenden Sekundärwand. Im letzten Bild werden bei höchster Vergrößerung die Durchmesser der Mikrofibrillen messbar. Im Rahmen der Messgenauigkeit ergeben sich bei den feinsten sichtbaren Strängen Durchmesser von ungefähr 10 nm. Die Größenordnung der Messwerte passt gut zu den Befunden von Giddings et al. (1980). Sie hatten festgestellt, dass bei der Formierung der Primärwand einzelne Produktionsapparate für Cellulose-Mikrofibrillen (die sogenannten Rosetten) Fasern mit einem Durchmesser von 5 nm produzieren. Sie konnten weiter zeigen, dass bei der Entstehung der Sekundärwand hexagonal angeordnete Gruppen von Rosetten Komplexfibrillen von 28 nm Durchmesser herstellen, welche die Basis der sekundären Wand bilden.

Abbildung 5: Detail der oberen Zellhälfte (Ausschnitte des Scheitellappens und der Seitenlappen). Das Maschenwerk der Cellulose-Mikrofibrillen der Primärwand sowie die Porenapparate (Pfeil) in der Sekundärwand werden klar erkennbar.



Abbildung 6: Vergrößerungsreihe (a-c), an deren Ende sogar die Dicke einzelner Netzstränge messbar wird. Die Pfeile weisen exemplarisch auf Porenapparate hin.
Solcherart instruktive Darstellungen des Primärwandgerüstes bei Zieralgen sind auch in der biowissenschaftlichen Literatur rar. Das wird wohl daran liegen, dass die ultrastrukturellen Details der Zellulosewände sowie die Prozesse um deren Entstehen mithilfe von Transmissionselektronenmikroskopen (TEMs) in den 1980er und 1990er Jahren erforscht worden sind. REMs in der Qualität, um solche Bilder wie oben gezeigt aufnehmen zu können, waren damals schlicht nicht verfügbar. Ein weiterer wichtiger Grund dürfte sein, dass dieses dünne Häutchen im Rahmen der Prozesse bei der Kritischpunkttrocknung üblicherweise verloren geht, was für die Darstellung der Oberflächenstruktur der Sekundärwand (deren Aussehen artbestimmungsrelevant ist) auch günstig ist.
4 Literatur
Bettighofer, W. (2025). Zellbiologische Beobachtungen an Desmidiaceen. Mitteilungsblatt der Mikroskopischen Gesellschaft Wien – Doppelheft 2&3, November 2025, 15–27.
Brook, A. J. (1981). The biology of desmids (Vol. 16). Univ of California Press.
Giddings Jr, T. H., Brower, D. L., & Staehelin, L. A. (1980). Visualization of particle complexes in the plasma membrane of Micrasterias denticulata associated with the formation of cellulose fibrils in primary and secondary cell walls. The Journal of cell biology, 84(2), 327–339.
Kleinig, H., & Maier, U. (1999). Zellbiologie: Begründet von Hans Kleinig und Peter Sitte.
Wanner, G. (2023). A practical guide to scanning electron microscopy in the biosciences. Wiley-VCH.